Article Text

Abstract
Background Many patients with pancreatic cancer (PC) suffer from abdominal pain and back pain. However, the cause of pain associated with PC is largely unclear. In this study, we tested the potential influence of the sonic hedgehog (sHH) signaling pathway on PC pain.
Methods Substance P (SP) and calcitonin gene-related peptide (CGRP) expression was measured in cultured PC cells and dorsal root ganglions (DRG) by real-time PCR, western blotting analysis and ELISA. Small interfering RNA transfection and plasmid constructs were used to regulate the expression of sHH in the AsPc-1 and Panc-1 cell lines. Pain-related behavior was observed in an orthotopic tumor model in nude mice.
Results In this study, the results show that sHH increased the expression of SP and CGRP in DRGs in a concentration and time-dependent manner. Additionally, sHH secretion from PC cells could activate the sHH signaling pathway and, in turn, increase the expression of nerve growth factor (NGF), P75, and TrkA in DRGs. Furthermore, the sHH signaling pathway and NGF/NGF receptor contributed to pain sensitivity in a nude mouse model.
Conclusion Our results demonstrate that PC pain originates from the sHH signaling pathway, and NGF mediates the pain mechanism via regulating SP and CGRP.
- pancreatic Cancer
- pain
- NGF
- sHH
This is an open access article distributed in accordance with the Creative Commons Attribution 4.0 Unported (CC BY 4.0) license, which permits others to copy, redistribute, remix, transform and build upon this work for any purpose, provided the original work is properly cited, a link to the licence is given, and indication of whether changes were made. See: https://creativecommons.org/licenses/by/4.0/.
Statistics from Altmetric.com
Introduction
Pancreatic cancer (PC) is one of the most aggressive malignant human cancers and is associated with an extremely poor prognosis.1 2 Patients with PC often experience abdominal and back pain.3 Many physiopathologic mechanisms are involved in PC pain, for example, nerve damage, inflammation, and perineural invasion (PNI).4–7 PNI is associated with poor survival, local recurrence, and neuropathic pain. Currently, pain is one of the major outcomes of the interaction between nerves and PC.8 9
Recent studies have demonstrated that expression of the sonic hedgehog (sHH) ligand contributes to PC progression.10 In addition, paracrine sHH signaling plays an important role in communication among the tumor, stroma, and nerves. sHH signaling is initiated by the binding of the sHH ligand to Patched-1 (PTCH1), which relieves repression of the transducer protein Smoothened (SMO), thus activating the GLi family of transcription factors. The activation of downstream factors such as GLi-1 and GLi-2 regulates target gene expression, resulting in proliferation, cell survival, and angiogenesis.11 As reported previously, sHH regulates nociceptive sensitization and guides the spatial pathfinding of raphespinal tract axons.12
Nerve growth factor (NGF) is a neurotrophic factor that belongs to the family of neurotrophins. NGF can bind tyrosine receptor kinases (Trk) and p75, a member of the tumor necrosis factor receptor superfamily, to promote neuronal survival and axonal growth. In the last decade, NGF and TrkA have been recognized as important mediators of pain.13–15 Additionally, NGF and TrkA are associated with the neuronal expression of inflammatory neuropeptides such as calcitonin gene-related peptide (CGRP) and substance P (SP).9 16
sHH signaling is directly involved in nerve cell activity and the regulation of PC progression.17 18 Some evidence strongly suggests that NGF and its receptors play a role in neuropathic pain.13 15 19 In this study, we demonstrate that PC pain originates from the sHH signaling pathway in nerves and that neurotrophic factors mediate the pain mechanism by regulating SP and CGRP.
Materials and methods
Orthotopic transplantation of tumor cells into nude mice
Prepared PC cells were injected into the pancreas of nude mice exposed by midline laparotomy (20 µL of cells at a total concentration of 1×106/μL, 4–6 sites; 20 µl total volume). After 4 weeks, the carcinoma model was successfully generated, and metastases to different locations did not occur by ultrasonic testing. Then, drug treatment and pain behavior could be tested.
Von Frey filament testing
After shaving the abdomens of nude mice, the abdomen of each mouse was stimulated 20 times at different intensities. Each stimulation lasted 2 s with an approximately 30 s interval between stimulations. A positive response consisted of lifting the belly and/or scratching or licking the abdomen. The number of responses was summed over all 20 stimulations. Each group had 10 mice for the testing.
Cell culture and reagents
The human PC cell lines AsPc-1 and Panc-1 were obtained from the American Type Culture Collection (Manassas, VA) and cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotics/antimycotics in a humidified 5% CO2 atmosphere at 37°C. Antibodies against sHH, SMO, PTCH, Gli1, Gli2, NGF, TrkA, p75, SP and CGRP were purchased from Abcam, USA. Recombinant sHH was obtained from R&D Systems (Minneapolis, MN).
Real-time PCR
Total RNA was extracted using TRIzol (Invitrogen, CA, USA), and cDNA was synthesized using a Prime Script RT Reagent Kit (TaKaRa, Dalian, China). The real-time experiments were conducted on an iQ5 Multicolor Real-Time PCR Detection System (Bio-Rad, Hercules, CA) using an SYBR Green Real-time PCR Master Mix (TaKaRa, CA, USA). The primers are shown in online supplementary table 1.
Supplemental material
Western blotting analysis
Cells were lysed using a lysis buffer (50 mM Tris (pH 7.5), 150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 1 mM EDTA, and 0.1% sodium dodecyl sulfate (SDS)) containing a protease inhibitor cocktail (Sigma-Aldrich), and protein concentrations were measured with the DC Protein Assay (Bio-Rad Laboratories, Hercules, CA, USA). After separation on 7.5% SDS-polyacrylamide gels, proteins were transferred to nitrocellulose membranes (Amersham Bioscience), which were then incubated with primary antibodies at 4°C overnight. After being washed three times with TBST, the membranes were incubated with horseradish peroxidase-conjugated secondary antibodies for 1 hour. Immunoreactive bands were visualized using an enhanced chemiluminescence kit (Millipore, MA, USA).
ELISA
To quantify the relative levels of pain factors in culture media and serum, an ELISA kit (R&D, Minneapolis, MN, USA) was used according to the manufacturer’s instructions.
Plasmid construction and transfections
Cells were seeded into small dishes and transfected with 100 nM siRNA using Lipofectamine RNAi MAX Reagent (Invitrogen, CA, USA) according to the manufacturer’s instructions. The cells were used for further experiments 24 hours after transfection. In addition, plasmids for the overexpression of sHH were constructed according to the manufacturer’s instructions to study the activity of the sHH signaling pathway.
Isolation and culture of mouse DRGs
DRGs were isolated from newborn rats, stored on ice in DMEM:F12 (1:1) containing 20% FBS, and then seeded into 24-well plates. The medium was changed every 3 days. DRGs were maintained at 37°C in a humidified atmosphere of 5% CO2.
Co-culture system for PC cells and DRGs
When the DRGs were isolated from newborn rats, DRGs were subsequently seeded on 24-well plates in 25 µL of an extracellular matrix (ECM) gel (BD Biosciences, Franklin Lakes, USA). The dishes were then placed in an incubator at 37°C with 5% CO2 for 30 min to enable the polymerization of the ECM gel, and then carefully submersed in 200 µL DMEM supplemented with 10% FBS. After 2–4 days, some axons from the DRGs cultured. Then the medium was changed, and PC cells (105/μL, total 50 µL) were directly placed exactly 2 mm from the DRGs. The co-cultures were incubated at 37°C with 5% CO2 in a humid atmosphere for 24 hours.
Drug treatments
In vitro, recombinant sHH was applied to DRGs at different concentrations for 12 hours. Cyclopamine, an sHH pathway inhibitor, was diluted to 18 µg/mL into DRG media for 12 hours. To detect the effect of neurotrophic factors, 60 ng/mL NGF was added into the DRG medium. GW441756 (15 µM for in vitro20, 30 mM for in vivo21), an inhibitor of TrkA, was used to block TrkA effects in DRGs by addition into the media and by treatment of animals by intraperitoneal administration.
Statistics
Analysis was performed using the SPSS statistical software package (V.13.0). The significance of the data was determined using Student’s t-test, one-way or two-way analysis of variance for in vitro and in vivo results. The significance level was set at p<0.05. All results are presented as the mean±SD.
Results
sHH increased SP and CGRP expression in DRGs
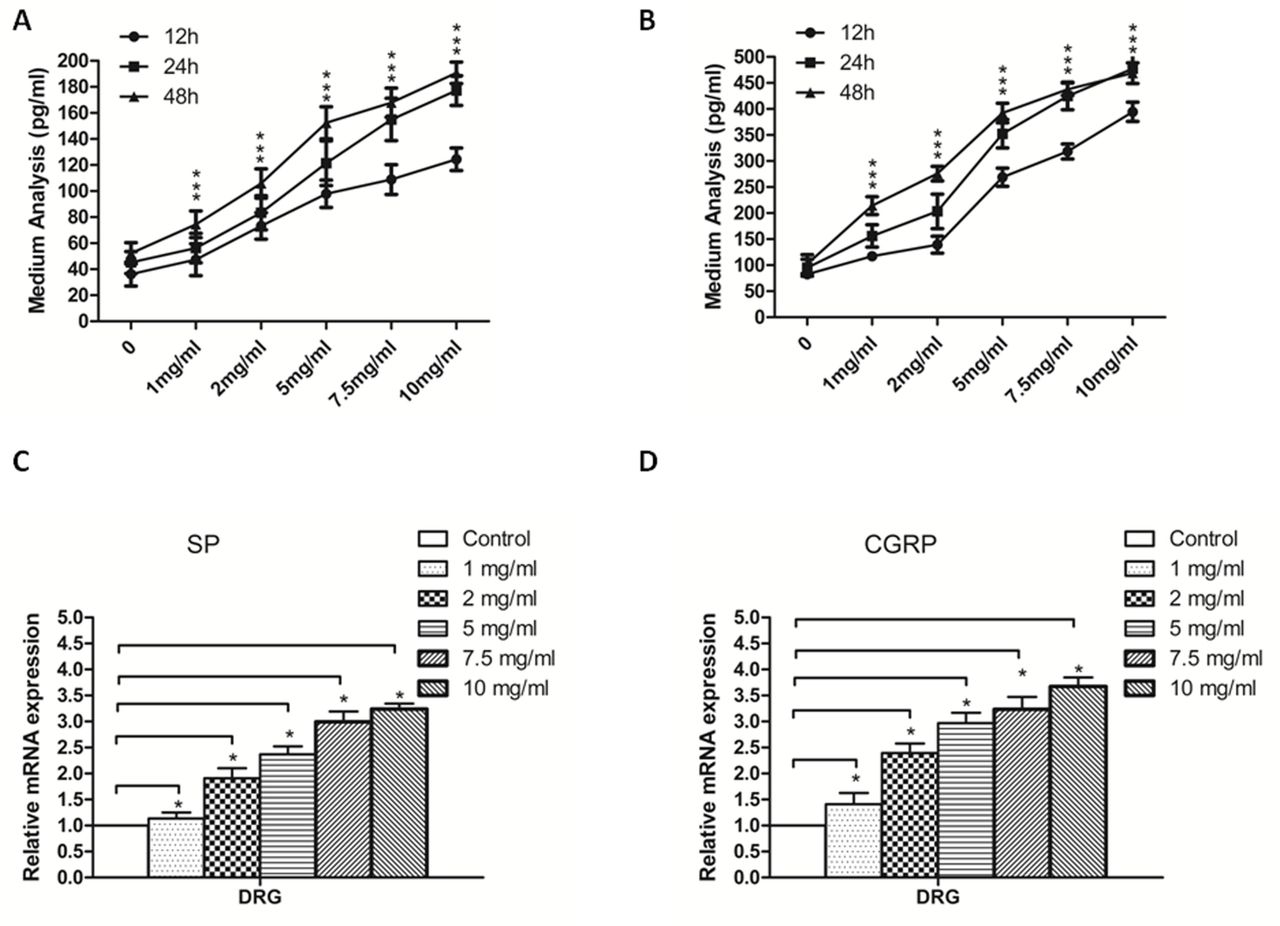
To investigate the possible relationship between sHH and pain factors in DRGs, we first evaluated how different concentrations of sHH affect DRGs. The mRNA and secretion levels of SP and CGRP in the culture medium were analyzed by ELISA and real-time PCR (figure 1). The results show that the secretion of SP and CGRP was dependent on the duration and concentration of sHH treatment (figure 1A,B) (p<0.05). Moreover, the mRNA results were similar to the ELISA results (figure 1C,D) (p<0.05). Thus, the primary dose–effect relationship between sHH and pain factors was detected. In the following experiments, sHH was used at 5 mg/mL for 24 hours.

Sonic hedgehog (sHH) increases the secretion and expression of SP and CGRP in DRGs. (A) SP secretion was analyzed in DRGs after treatment with different concentrations of sHH at 12, 24, and 48 hours. (B) CGRP secretion was analyzed in DRGs treated with different concentrations of sHH at 12, 24, and 48 hours. (C) SP mRNA expression was determined in DRGs treated with different concentrations of sHH at 24 hours. (D) CGRP mRNA expression was determined in DRGs treated with different concentrations of sHH at 24 hours. The results showed that the secretion of SP and CGRP was dependent on the concentration and timing of sHH. *P<0.05 compared with control. All data from three independent experiments were analyzed. CGRP, calcitonin gene-related peptide; DRG, dorsal root ganglion; SP, substance P.
Co-culture of sHH-expressing DRGs and PC cells increased SP and CGRP expression
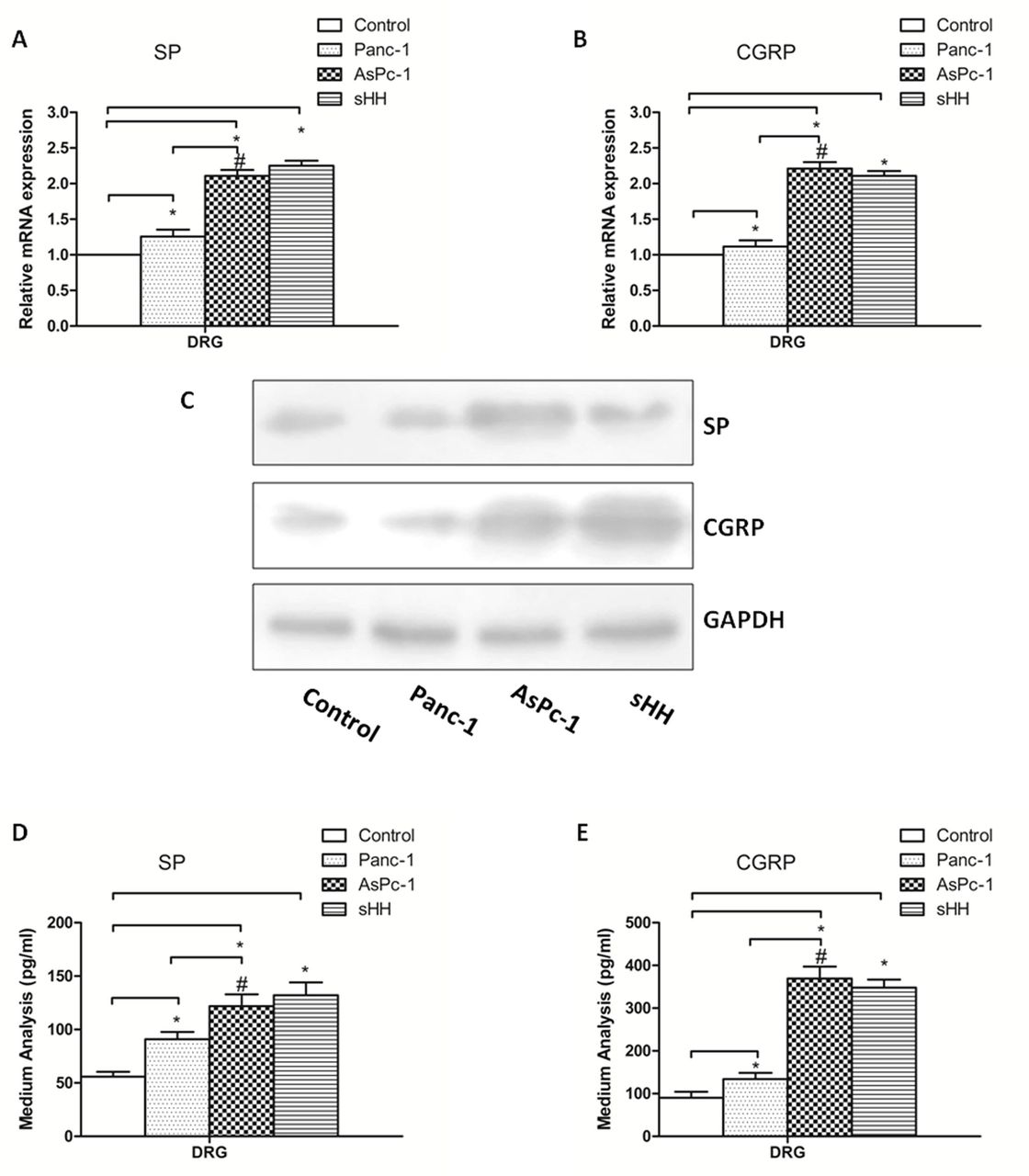
To determine whether PC cells mediate pancreatic pain by sHH, Panc-1 (low sHH expression) and AsPc-1 cells (high sHH expression) were used in our study. The expression and secretion of SP and CGRP were measured by ELISA, real-time PCR and western blotting in the co-culture system (PC cells were directly co-cultured with DRGs for 24 hours) (figure 2).
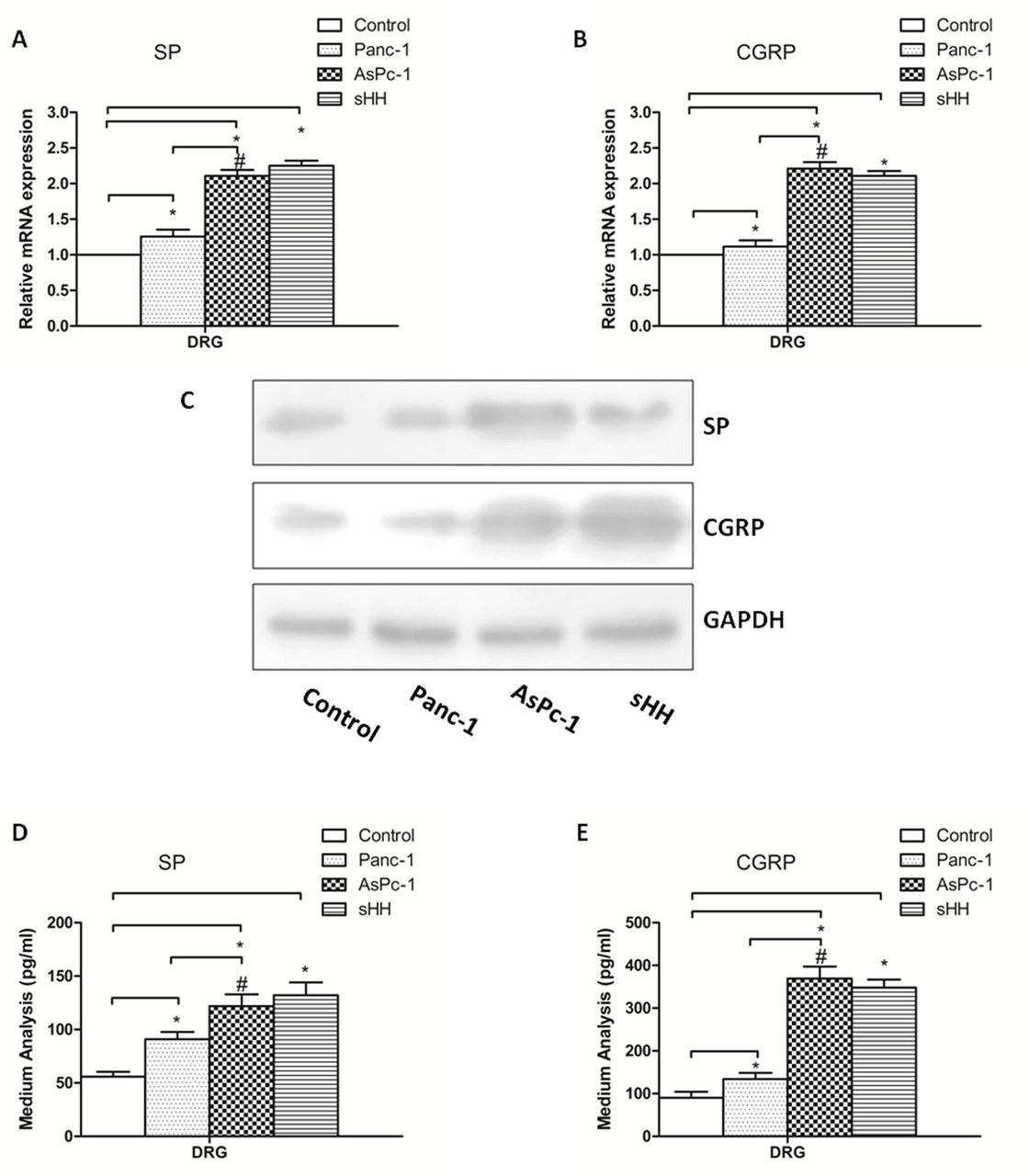
Co-culturing pancreatic cancer (PC) cells and DRGs increases secretion and expression of SP and CGRP in DRGs. (A, B) The mRNA expression of SP and CGRP was higher in AsPc-1 cells (high sHH expression) than in control and Panc-1 cells (low sHH expression). (C) The protein expression of SP and CGRP was regulated by PC cells. (D, E) Secretion levels of SP and CGRP were regulated by PC cells in DRGs. The results showed that protein expression and secretion results were similar to mRNA results. *P<0.05 compared with control. #P<0.05 between AsPc-1 and Panc-1 cells. All data from three independent experiments were analyzed. CGRP, calcitonin gene-related peptide; DRG, dorsal root ganglion; sHH, sonic hedgehog; SP, substance P.
The mRNA results showed that the expression levels of SP and CGRP in DRGs were much higher in the co-cultured PC cells and sHH group than in the control group (figure 2A,B) (p<0.05). Additionally, there was a significant difference between the Panc-1 and AsPc-1 co-culture groups (p<0.05). To supplement the mRNA analyses, western blotting was used to quantify protein levels, and the results showed that protein expression was similar to mRNA expression (figure 2C) (p<0.05). Additionally, the concentrations of SP and CGRP in the culture medium were analyzed by ELISA. The results showed that the secretion of pain factors was much greater in the co-culture and sHH groups (figure 2D,E) (p<0.05).
sHH from PC cells activated the sHH signaling pathway in DRGs
To determine whether sHH secretion from PC cells could activate the sHH signaling pathway in DRGs, the PC cell lines Panc-1 (low sHH expression) and AsPc-1 (high sHH expression) were transfected (figure 3). The transfection efficiency was checked by real-time PCR and western blotting (figure 3A–D). Moreover, we measured the expression of sHH pathway signaling molecules in DRGs by western blotting in the co-culture system (figure 3E). The results showed that overexpressing sHH in Panc-1 cells induced the expression of the transcription factors Gli1 and Gli2 compared with control and cyclopamine-treated cells (p<0.05). In addition, knockdown of sHH in AsPc-1 cells using siRNA inhibited the expression of target molecules (p<0.05).

si-sHH-transfected pancreatic cancer (PC) cells activate the sHH signaling pathway in dorsal root ganglions (DRG). (A, B) The transfection efficiencies were verified by western blotting. Panc-1 cells (low sHH expression) were transfected with sHH plasmids, and AsPc-1 cells (high sHH expression) were transfected with sHH siRNA. (C, D) The transfection efficiencies were verified by real-time PCR. (E) sHH pathway signaling molecules in DRGs were checked by western blotting. The results showed that overexpressing sHH in Panc-1 cells induced the expression of the transcription factors Gli1 and Gli2, and knockdown of sHH in AsPc-1 cells using siRNA inhibited the expression of target molecules. *P<0.05 compared with control. sHH, sonic hedgehog.
Activation of the sHH signaling pathway increased the expression of NGF, P75, and TrkA in DRGs
To determine whether the activation of the sHH signaling pathway significantly contributes to the induction of NGF and receptors in DRGs, we measured the expression of NGF, P75, and TrkA in the co-culture system using real-time PCR and western blotting (figure 4). The mRNA results showed that NGF, P75, and TrkA were increased in DRGs co-cultured with sHH-overexpressing PC cell lines. Furthermore, the expression of NGF, P75, and TrkA could be reversed by treatment with cyclopamine or sHH siRNA (figure 4A,B) (p<0.05). Similar results were obtained with PCR and protein analysis by western blotting (figure 4C). Thus, activation of the sHH signaling pathway induced the expression of NGF and receptors in DRGs.

sHH signaling pathway activity increases the expression of NGF, P75 and TrkA in dorsal root ganglions (DRG). (A, B) The expression of NGF, P75 and TrkA in DRGs was checked by real-time PCR. The results demonstrated that NGF, P75 and TrkA were increased in DRGs co-cultured with pancreatic cancer (PC) cell lines that overexpress sHH. The expression of these molecules could be reversed by cyclopamine or siRNA against sHH. (C) The expression of NGF, P75 and TrkA in DRGs was checked by western blotting. *P<0.05 compared with control. NGF, nerve growth factor; sHH, sonic hedgehog.
sHH increased SP and CGRP expression in DRGs in an NGF-dependent manner
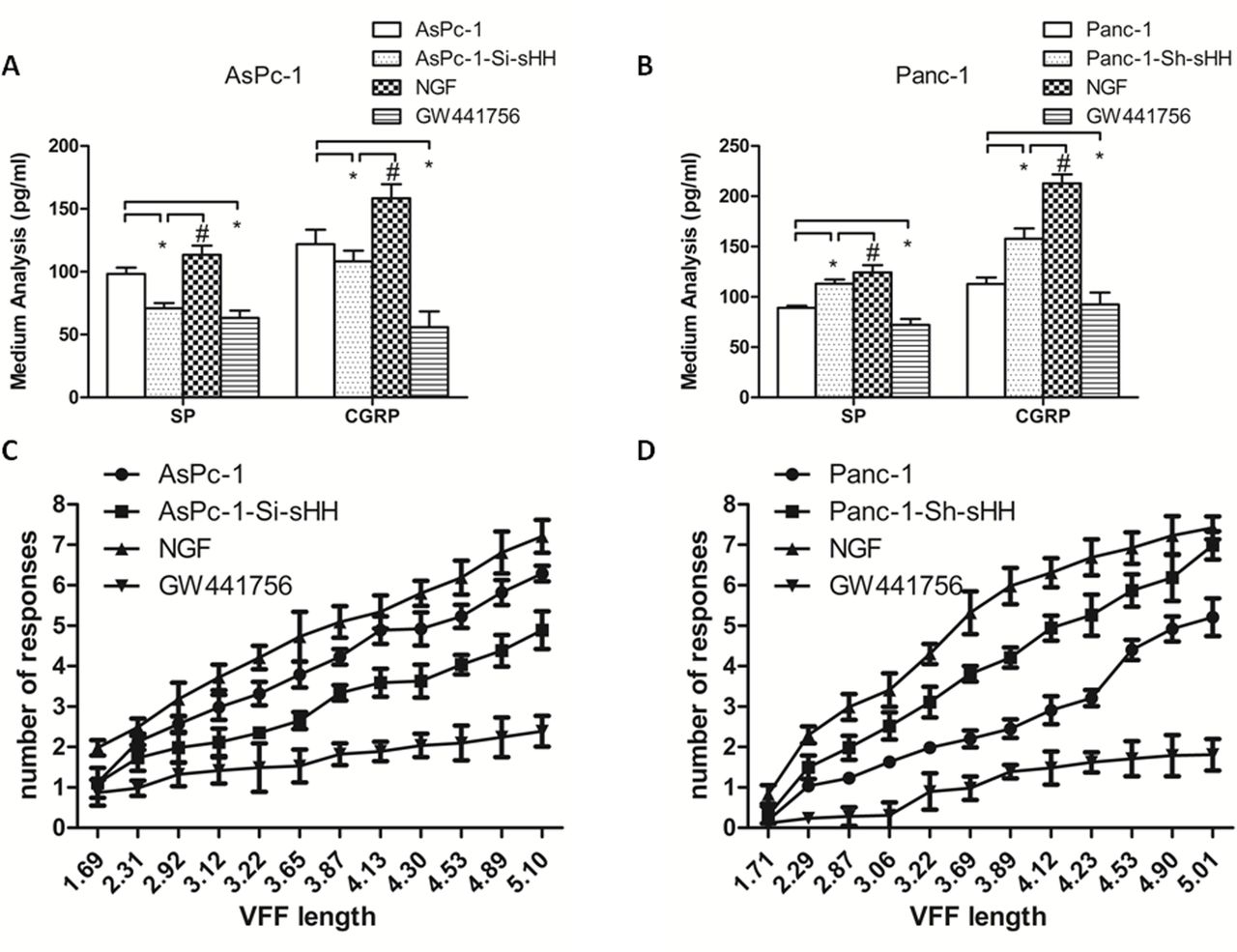
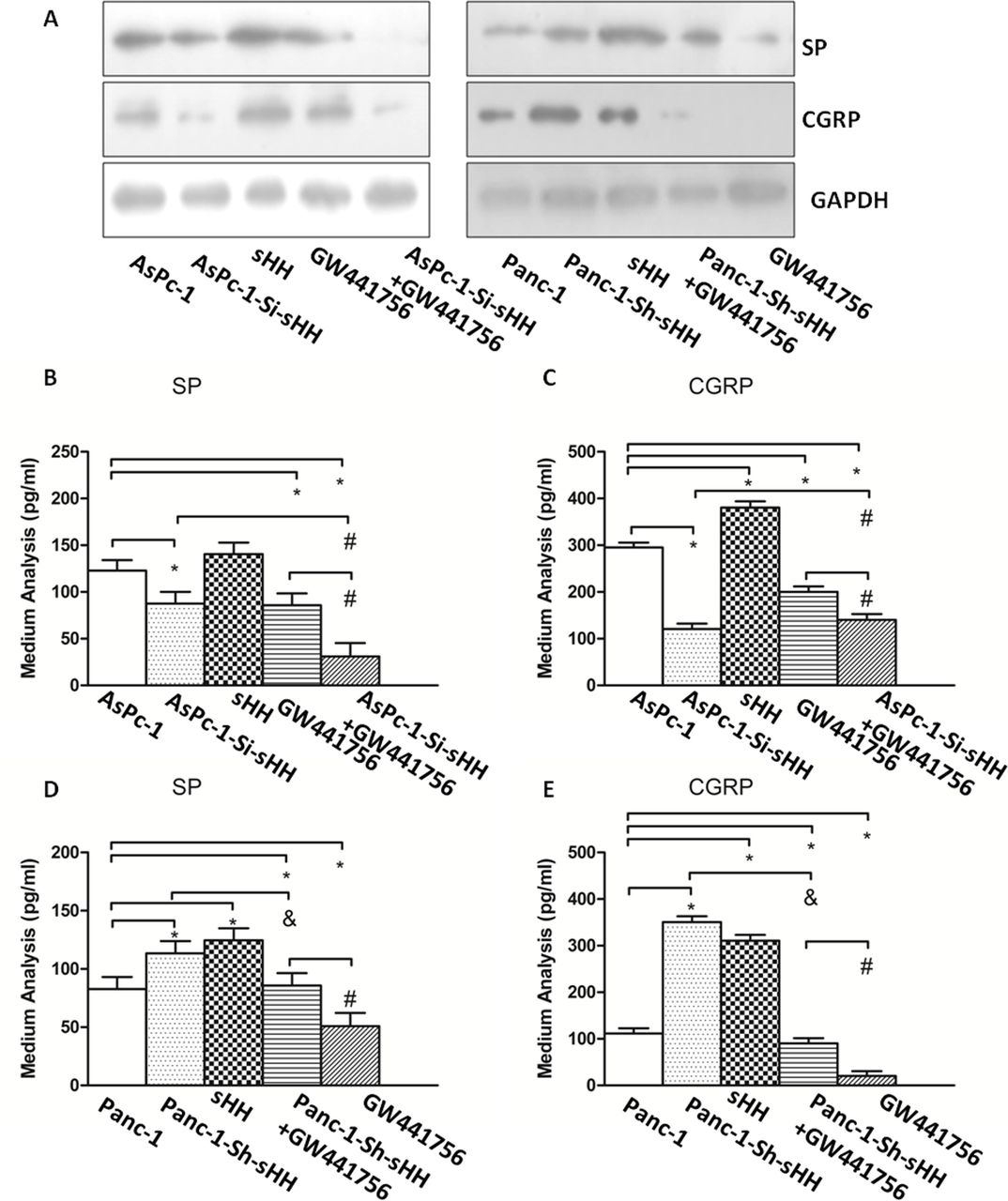
To determine whether NGF contributed to the expression and secretion of SP and CGRP in the co-culture system, we quantified the expression levels of SP and CGRP in DRGs and in the medium by western blotting and ELISA, depending on the different groups (figure 5). The results showed that treatment with GW441756, an inhibitor of TrkA, alone or in combination with cyclopamine diminished the expression of SP and CGRP compared with control (figure 5A) (p<0.05).

Nerve growth factor (NGF) and sHH increase the expression and secretion of SP and CGRP in dorsal root ganglions (DRG). (A) The expression levels of SP and CGRP were checked by western blotting. The results showed that GW441756, alone or in combination with cyclopamine, diminished the expression of SP and CGRP. (B, C) The secretion of SP and CGRP was determined by ELISA in the co-culture system containing AsPc-1 cells and DRGs. *P<0.05 compared with control. #P<0.05 compared with AsPc-1-Si-sHH+GW441756. (D, E) The secretion of SP and CGRP was determined by ELISA in the co-culture system containing Panc-1 cells and DRGs. *P<0.05 compared with control. #P<0.05 between GW441756 and Panc-1-Sh-sHH+GW441756. &P<0.05 between Panc-1-Sh-sHH and Panc-1-Sh-sHH+GW441756. All data from three independent experiments were analyzed. CGRP, calcitonin gene-related peptide; sHH, sonic hedgehog; SP, substance P.
The results showed the greatest inhibitory action in the combined group in AsPc-1 cells (figure 5B,C) (p<0.05). However, there was no difference between AsPc-1 cells transfected with si-sHH and those treated with GW441756 (p>0.05). Additionally, in Panc-1 cells, the inhibitory effect of GW441756 was stronger than that of the other treatments/conditions (figure 5D,E) (p<0.05).
sHH and NGF/NGF receptor determined pain sensitivity in a nude mouse model
To determine whether sHH and NGF/NGF receptor mediate PC-associated pain behavior in a mouse model, we designed an in situ tumor model in nude mice. PC cells were injected into the pancreas of nude mice. After 4 weeks, we measured the sensitivity of the abdomen to mechanical stimulation by the von Frey filament probing method. In addition, we quantified the secretion of SP and CGRP in the sera of each group of mice 2 hours after mechanical stimulation (figure 6). The frequency of response to mechanical stimulation for mice injected with NGF was greatest, and injection with GW441756 decreased the response to stimulation (figure 6C,D) (n=10) (p<0.05). Panc-1 cells transfected with sh-sHH led to a greater sensitivity than Panc-1 control cells, and AsPc-1 cells transfected with si-sHH led to weaker sensitivity than those treated with NGF control. ELISA analysis revealed that the concentrations of SP and CGRP were consistent with the pain behavior results (figure 6A,B) (p<0.05).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Sonic hedgehog (sHH) and NGF aggravate pain behaviors and induce the secretion of SP and CGRP in mice. (A, B) NGF treatment and overexpression of sHH in pancreatic cancer (PC) cells resulted in greater pain sensitivity and higher secretion of SP and CGRP. However, GW441756 treatment and downregulation of sHH in PC cells decreased the secretion of SP and CGRP in the co-culture system. *P<0.05 compared with control. #P<0.05 between NGF and AsPc-1-Si-sHH group (for A); or between NGF and Panc-1-Sh-sHH (for B). (C, D) The response rate was the greatest in the NGF group. The weakest response was observed in the AsPc-1-si-sHH group. All data from three independent experiments were analyzed. CGRP, calcitonin gene-related peptide; NGF, nerve growth factor; SP, substance P; VFF, von frey filament.
Discussion
PC is one of the most aggressive solid tumors, and sHH levels are elevated in human PC compared with normal pancreatic tissue.22 PNI extending into the extrapancreatic nerve plexus is a histopathological characteristic of PC and leads to abdominal pain.6 NGF induces thermal and mechanical sensitization of neurons, and the recent use of NGF-sequestering antibodies attests to the ongoing role of this neurotrophin in chronic pain.23 24 However, the specific mechanism contributing to PC pain is unclear.
Previous studies have found that sHH is abnormally expressed in PC and is associated with cancer development and progression.10 Furthermore, inhibition of sHH signaling is an attractive clinical target for therapeutic intervention.25 In addition, our preliminary experiment demonstrated that the sHH signaling pathway contributed to PNI and PC pain by pancreatic stellate cells (PSC).26 In the current study, we first showed that the expression and secretion of SP and CGRP were dependent on the timing and concentration of sHH activity in DRGs. Then, we explored sHH expression in PC cells, where this ligand acts in a paracrine manner on DRGs to increase activation of sHH signaling. This is a new model of cancer–nerve interaction involving sHH, nerve cells, and NGF in PC pain. In this study, we also showed that sHH increased the expression of NGF in an sHH-dependent manner in DRGs. The results demonstrated that NGF, P75, and TrkA were increased in DRGs co-cultured with sHH-overexpressing PC cell lines. Additionally, the increased expression of NGF, P75, and TrkA could be reversed by treatment with cyclopamine or si-sHH.
Recent studies have suggested that the tumor microenvironment plays a considerable role in cancer cell proliferation, malignant progression, and response to therapy.27 Therefore, the interaction between the tumor stroma and PC cells may represent a novel therapeutic option for PC.28 Indeed, our previous results showed that PSCs played an essential role in the pain mechanism in PC.26 Briefly, sHH secreted from PC cells activates the sHH signaling pathway in PSCs and enables the complex pain process via NGF and brain-derived neurotrophic factor upregulation or sensitization to TRPV1 and pain factors in sensory nerves.
In previous studies,26 we discovered that PSCs contribute to neuropathic pain but do not play an indispensable role in the mechanism. Thus, we focused on the direct relationship between PC cells and DRGs through the sHH signaling pathway. The results confirmed that our hypothesis was feasible and that NGF was involved in neuropathic pain. The results also showed that an inhibitor of TrkA, alone or combined with cyclopamine, diminished the expression of SP and CGRP.
The mechanical interactions between the tumor microenvironment and cancer pain are complicated and involve multiple factors. The sHH signaling pathway is not the only factor connecting PC cells and DRGs to contribute to neuropathic pain. Thus, despite the limitations of our research, this work contributes to the understanding of cancer-related pain and highlights areas that require further study.
In addition, the DRG was regarded as a whole in this study, and we did not differentiate the cell components. Schwann cells (SC), which are equally present in visceral nerves, were exposed to the hallmark components of the PC microenvironment.29 30 SC activation could decrease pain sensation in patients with PC and decrease mechanical hyperalgesia in mice with PC due to concomitant suppression of spinal astrocytes and microglia.29 Further follow-up experiments are needed to clarify the detailed mechanism of PC pain.
NGF is known to activate and promote the growth of sensory neurons. NGF and its interaction with its cognate receptor TrkA, a common marker of peptidergic neurons together with CGRP in the DRG, have come to be recognized as important mediators of pain.19 Our study provides evidence that NGF plays an important role in PC pain through regulation of sHH signaling. Thus, these findings suggest that NGF and TrkA are important for the generation and control of neuropathic pain.
In this study, an orthotopic tumor model was used to examine the experimental hypothesis in vivo. Von Frey filament testing31 showed a significant increase in pain behavior in the NGF and sHH groups. The results demonstrated that sHH signaling and NGF are involved in pain generation in the animal model, consistent with the results of our previous in vitro studies.26 However, the mechanism of neuropathic pain is complex, and many other elements may be involved. Whether the SP and CGRP changes are mediated by different factors remains unknown. Thus, our results illuminate a path for further study.
Conclusions
In conclusion, sHH secreted from PC cells can increase the expression of NGF and TrkA by activating the sHH signaling pathway in DRGs. Additionally, the downstream pain factors induce expression changes in response to sHH and NGF. Our findings strengthen the rationale for PC-associated pain generation and may provide evidence for a novel therapeutic option for cancer pain.
References
Footnotes
LH and JJ are joint first authors.
Contributors Design: LH and QM. Writing: LH and JM. Analysis: JJ. Methodology: TQ, YX and MX. Data curation: EW and XS. All authors read and approved the final manuscript.
Funding This study was supported by the National Natural Science Foundation of China (81502074, 81672434, 81702916).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The Ethical Committee of the First Affiliated Hospital of Xi’an Jiaotong University.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available in a public, open access repository. There are no data in this work. Data are available upon reasonable request. Data may be obtained from a third party and are not publicly available. All data relevant to the study are included in the article or uploaded as supplementary information.
Author note Corresponding author:
Dr Qingyong Ma, Department
of Hepatobiliary Surgery, Xi’an
Jiaotong University Medical
College First Affiliated Hospital,
Xi’an, China;qyma56@ xjtu. edu. cn;
Dr Jiguang Ma, Department of Anesthesiology,Xi’an
Jiaotong University Medical
College First Affiliated Hospital,
Xi’an, China;
jgma86{at}mail.xjtu.edu.cn